
Brief Introduction to Cephalopod Biology:
Cephalopods are a class of marine mollusks which include octopods, squid, cuttlefish and the nautilus. The Greek ‘Cephalopoda‘ translates to ‘head-foot’; a fitting description of the characteristic anatomical features of these peculiar molluscan organisms. Scientists from many disciplines have begun to pay closer attention to cephalopods because of their extraordinary neurological, anatomical, physiological, behavioral and adaptive qualities. In this article, I will attempt to provide the reader with a summary of the reasons why cephalopods have grown in popularity among scientists and the public alike.
In order to properly understand cephalopod biology; we must first review a little bit about what it means to be a mollusk.

Molluscan’s represent among the most extravagant examples of adaptive radiation in the whole of Evolutionary Zoology; with seemingly endless varieties of mollusks all sharing a simple body plan as a result of common ancestry. The molluscan shell may be bivalve (having 2 lateral, jointed plates, e.g. clams), univalve (single shelled, e.g. snails), plated (e.g. Polyplacophoran’s) or internal (e.g. cephalopods, slugs, sea hares). Some classes of mollusk have distinct male/female individuals which engage in sexual reproduction, others are hermaphroditic. One size extreme among mollusks is represented by giant marine squid (family Architeuthidae), which may achieve an adult size of up to 45 feet long. Below is a photograph of the dissection of one of these massive creatures.

The other size extreme is represented by the so-called micro-mollusks below, which achieve an adult size of only 5-6 mm.

Being that we are terrestrial organisms, many of us make the mistake of assuming a generalized mollusk would be something like a snail. On its face, this seems an easy enough mistake to make considering the variety of snails we see in our gardens, near streams and even at the beach. However, common though they are, many mollusks have features quite strikingly different than those of the snails. Cephalopods, Sea Hares and Slugs are a few examples of molluscan’s who tip the idea of a snail being a ‘generalized mollusk’ on its head. These mollusk groups have internal shells which are relatively small compared to their body size, while other mollusks like snails and clams have external shells which are large relative to their body size. Some mollusks use an internal means of fertilization, others use external fertilization. For these reasons, it can become difficult to provide a description of a ‘generalized mollusk’.
Some wonder how a ‘molluscan family reunion’ could include members as diverse as the giant squid and the micro mollusks. Despite my personal reluctance to accept the notion of a ‘generalized mollusk model’, it can certainly be said that some characteristics are shared among all mollusks. One of such features is the presence of a molluscan mantle, which secretes the material necessary for shell formation and is where the kidneys and anus open. Depending on the species of mollusk, the position of the mantle can be quite different (see hermaphroditic snail anatomy below).
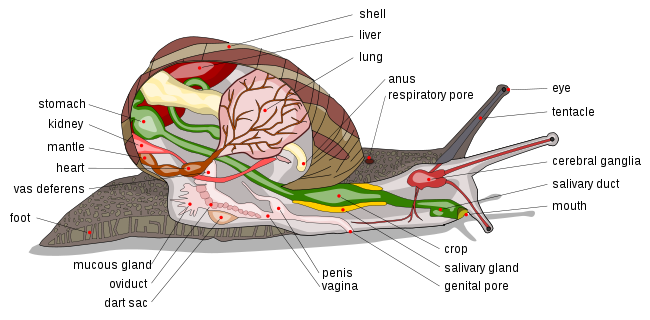
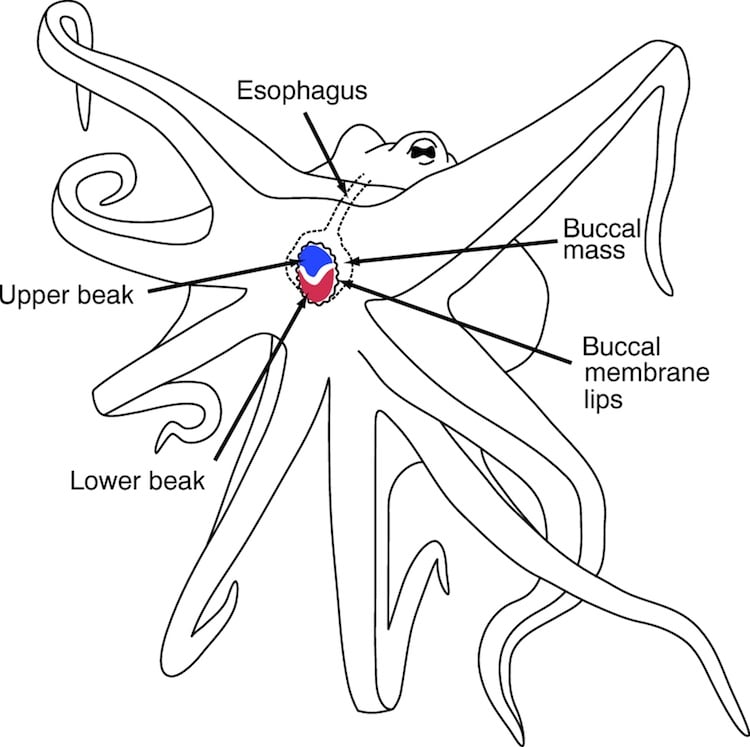
Mollusks also have a muscular region called a ‘foot’, which has a locomotive function. Pairs of sensors (called statocysts) provide mollusks with their sense of balance and them help maintain equilibrium with respect to gravity; serving a similar purpose to that of the inner ear region in humans. Another similarity among mollusks is the ganglionic (peripheral, compartmentalized) nature of their nervous systems. Not all species of mollusk have centrally located ‘brains’; but in those species that do, the brain forms a sort of doughnut around the animals esophagus (muscular tube through which food passes). The alimentary canal (digestive pathway, from entrance to exit) of mollusks is often connected to a region called a buccal mass, see below.
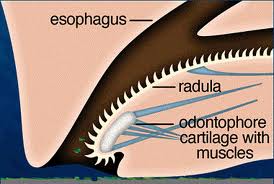
Additionally, mollusks have ‘open’ circulatory systems, meaning that their blood is pumped into a hemocoel, where blood diffuses back into circulatory system between individual cells. This is quite different than the closed circulatory systems of we humans and other vertebrates; which are composed of a heart pumping blood (that does not usually fill body cavities) through a series of blood vessels of varying size and width. Mollusks respire using paired gills called ctenidia, which divide the mantle cavity and are usually feather-shaped and capable of gathering oxygen from the organisms external environment. Mollusk ‘tongues’, called radulae, are composed of many rows of chitin-based ‘teeth’ which are used to break down their food into manageable sized pieces prior to digestion (see radula illustration below).
…Now that we have reviewed the curious biological features of the enormously varied phylum Mollusca, I will begin to narrow my focus to Cephalopod studies; more specifically, the study of octopods…
Why are Octopods interesting, Anatomically/Physiologically?
Octopod Eyes –
Based on all the assessments of the eye of the cephalopod, they really are color blind, in their eyes; so how can they adapt with such perfect accuracy to the color of their background? A couple interesting features of cephalopods allow this to be possible. Cephalopods have skin cells which allow light from their environment to be reflected, giving off similar hues even if the cephalopod couldn’t detect the color themselves. But Roger Hanlon of the Marine Biological Laboratory in Massachusetts has proposed another possibility. The active genes coding for an opsin molecule often associated for color vision in vertebrates was found in the skin of an octopus’ arm; suggesting that distributed ganglia within the arms may be able to detect the color of the animals environment. If this is the case, such information would be relayed back to the animals central brain where an overall image of its environment is assembled from the information received from the eyes and distributed ganglia.
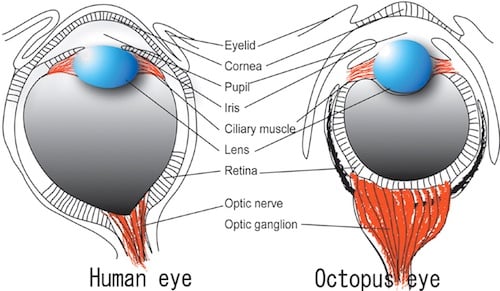
The unique formation of the octopus eye has had the attention of many scientists; in fact, many view its natural ‘design’ to be superior to the ‘design’ of our own eye. An excellent analogy given by Dr. Randolph Nesse sums up the issues with our vision: “Imagine a camera designer who put wires in between the light and the film, that is how our eye works. Not only that, our eye has a whole blindspot where nothing can be seen”. In light of this information, biological engineers have joked that if they were ‘designing’ us, and had been given a blueprint of a human eye, they would have sent it back for revision. To make up for this error our eyes, and indeed the eyes of all other vertebrates, constantly jiggle a bit in the socket. This jiggling, called nystagmus, means the area of vision is never in the exact same position; allowing the brain to form a comprehensive picture of the organisms environment. Because of this corrective mechanism, we are not able to notice that we have a blind spot, though we most certainly do.
The evolutionary history of humans and cephalopods is quite drastically different, with our lineages having separated over 500 million years ago. The differences in the eye of the human and the octopus are quite extensive; and octopods don’t have many of the previously mentioned issues with vision that we humans have. But why not? This mainly has to do with the genetic and developmental differences between our species. Unlike in humans, during octopus development they eyes develop with the vessels and nerves exiting from the back of the eyeball. This developmental difference seems small- yet it is responsible for the fact that octopods, unlike vertebrates, can NOT get retinal detachments, never have a problem with a blind spot and do not require constant eye jiggling to attain a complete field of vision.

Octopod Arms and Suckers:
As most of us are aware, octopods have eight arms which are covered in suckers. Depending on the octopus species, each arm may have up to 300 individual suckers. Suckers often occur in pairs, and each is capable of tasting and smelling its environment. These bizarre suckers may work alone or coordinate with one another; in which case, suckers located across from (and next to) one another can pass bits all the way to the centrally located mouth. In some cephalopod species, these suckers are lined with tiny hooks or ‘teeth’ that enable them to have an extremely powerful grip of their prey items.
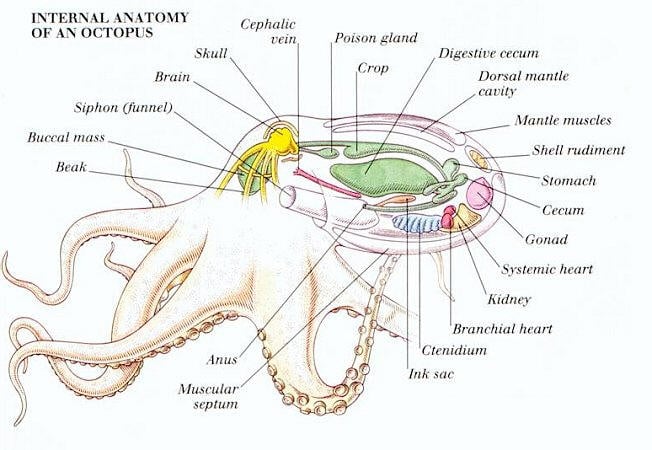
Octopod Hearts and Circulation:
Octopuses have three hearts and blue blood, which are difficult concepts for some to grasp. A possible explanation for their requirement for three hearts is that the molecule they utilize to transport oxygen (hemocyanin) is not as effective at this as the molecule (hemoglobin) used by vertebrates. Hemoglobin contains iron and, in short, its a more effective, complex oxygen transporting molecule than hemocyanin – which does not contain iron. When the iron in hemoglobin binds with oxygen, the oxygenated blood turns red. However, our blood (and the blood of other vertebrates) is also blue when it is depleted of oxygen. If you look at the veins on your wrist, which are pumping oxygen-depleted blood towards the heart, you will notice the blood they contain is also blue. Two of the cephalopods hearts are called ‘brachial’ hearts, and are responsible for pumping blood through the animals gills to oxygenate it. The other, larger heart, called the ‘systemic’ heart, is responsible for the circulation of blood throughout the organisms entire body.

Octopod Beak and Radulae:
The mouth of the octopod is a crushing, shredding machine; which effectively grinds the unfortunate prey animal into digestible portions in very little time. Such prey would first be met by the animals extremely powerful (and somewhat bird-like) beak, pictured above. This beak is very sharp and able to slice manageable chunks of the prey which then move on to the next step of processing. Next, the larger chunks of food are ground into what one would imagine to look similar to hamburger meat using the sharp spikes of this fearsome predators muscular radula, pictured below (which moves back and forth, continually shredding the food). After the food is sufficiently ground, the animal uses its muscular esophagus to force the food down its alimentary canal for further digestion.

Why are Octopods interesting, Neurologically?
Octopods and their kin have long been largely misunderstood by human, even in enlightened intellectual circles. Science has begun to take a different approach to the research many species of animals, and are finding extraordinary examples of complexity regarding animal behaviors and cognition; often in animals (like octopods) that many had previously written off as simple, thoughtless creatures. Even one of the most celebrated minds in the whole of human history, Aristotle, was convinced that octopods were stupid, as is referenced by the following quote:
“The octopus is a stupid creature, for it will approach a man’s hand if it be lowered in the water; but it is neat and thrifty in its habits: that is, it lays up stores in its nest, and, after eating up all that is eatable, it ejects the shells and sheaths of crabs and shell-fish, and the skeletons of little fishes”.

On one hand, Aristotle had a point. After all, if you put your hand in the water, you wouldn’t expect a fish to swim into it; thereby allowing an easy capture and a meal to the human. For this reason, it is understandable how one could see such a behavior as being ‘stupid’ from our perspective. However, this view fails to take into consideration that the octopus has absolutely no idea what the humans intentions are; it has probably never seen one before. The fact that octopods may approach a human, even interacting with them at will occasionally, doesn’t demonstrate stupidity as much as it does curiosity. In my experience working with animal behavior, curiosity is often an indicator of complex cognitive function (e.g. Primates, Parrots, Dogs, Cats).

Taking one question into more serious consideration sheds more light on the matter of cephalopod cognition: What all does octopus have to do to get away from predator? The answer is quite a lot, and all of it almost simultaneously. If a shark were to spot an octopus, the frightened animal would need to take into consideration where the predator is, where the hiding spots are, how much ink to squirt and in which direction, and what color / texture it needs to become to camouflage itself against the background – all of this in the matter of a couple seconds, or less. This demonstrates a wide awareness of the organisms surroundings and a complex capacity to process vast amounts of information simultaneously. How is this possible, do they have enormous brains?
The question about the animals brain comes up quite often, and the answer is a bit complicated. In short, octopods DO have large brains – larger than that of any other invertebrate and, relative to its body size, well up the mammalian brain size scale as well. However, their nervous systems function in a manner that is difficult for us to relate to or understand. The peripheral nervous systems of cephalopods are composed of many clusters of distributed neural ganglia; which are able to process information locally and send data back to the central brain. The ganglia send back information to the centralized brain and allow for the speeding up of motor processing.
In fact, approximately 2/3 of the octopuses average of 500 million neurons are located in the animals arms. About 50-75 million neurons are associated with the animals central brain, 130-150 million with the large optic lobes and all the rest are distributed in clusters found all over the animals body. While ganglia are composed of nervous tissue that looks very similar to the kinds of tissue we find in the animals central brain, the ganglia are all under the control of the animals central brain. The central brains responsibility is to form a cohesive image of the octopods environment from the data obtained from the animals peripheral senses.
Invertebrates without a complex, centrally located brain do not function this way. For example, if you were to cut a lobster in half, you would more or less make two (extremely uncomfortable) lobsters, which would die shortly thereafter. Because an octopus, like humans, has a complex, centrally located brain which constructs an image of our environment from information received from our various sensory organs – if we were to cut one in half (horizontally), the half connected to the brain would feel pain, but the part of the body no longer connected to the brain would not be capable of ‘feeling’ anything.
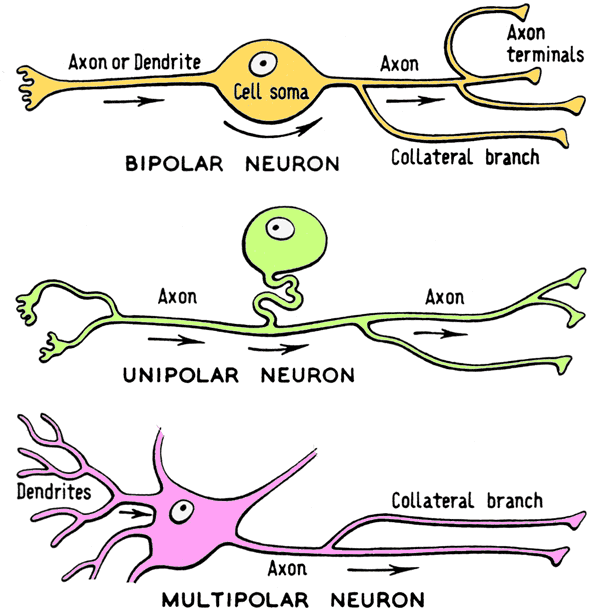
So, on one hand we are talking about a complex and dense brain that has tissue somewhat similar to complex vertebrate nervous tissue, but on the other hand, we see patches of distributive ganglia which are quite unlike anything we vertebrates are used to. Are there any important differences between the individual neurons of vertebrates and invertebrates?
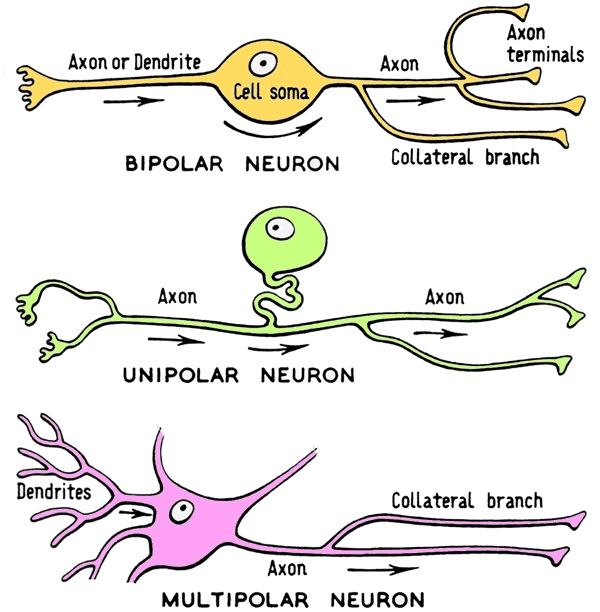
The answer is yes. Humans have more complex, multi-polar neurons; whereas most invertebrates have simpler unipolar neurons. The unipolar neurons of octopods lack an attribute seen in the cells of vertebrate animals called a myelin sheath. The myelin sheath acts an insulator of the electro-chemical impulses between nerves, which allow for faster transfer of information. Being that cephalopods have very quick reaction times and no myelin sheath, how is this possible? Some have solved this problem by evolving with very large axons, the size of which allow for direct, fast transmission of electrochemical impulses. Some cephalopod axons are so large, they can be seen without a microscope. Additionally, because invertebrates have bodies covered in peripheral ganglia, information is able to be sent back to the central brain in a way that is very different, but equally effective, to the neuro-communicative approach taken by vertebrates.
Interestingly, neuroscientists have also found functional similarities between the vertebrate hippocampus, associated with learning and memory, and the vertical lobes of the octopus brain. If an octopus has its vertical lobe damaged, it can no longer learn new mazes, problems or behaviors. It may be able to fumble through mazes it had learned in the past, but it will not be able to learn new ones. This is somewhat similar to what would be expected of a human who suffered an injury to their hippocampus. These similarities are examples of convergent evolution, octopods and humans haven’t shared a common ancestor in over 500 million years; therefore, if there are similarities to the way we perceive and process information, we don’t have these homologous characteristics as a result of common ancestry – rather these features arose independently of one another in humans and octopods because thats just how intelligence/complexity work.
Why are Octopods interesting, Behaviorally?
Octopods have been the focus of some quite fascinating cognitive research, and have shown their ability to possess both short and long term memory, navigate complex mazes, recognize color/pattern sequences and, controversially, even use tools and learn by observation. Claims of observational learning in baby cuttlefish were made from a marine biology laboratory in France. In this experiment, baby cuttlefish that were still developing in their translucent eggs were exposed to tanks including different kinds of prey. Generally, the only thing you can get baby cuttlefish to eat in captivity is small shrimp. However, clutches of these eggs were hatched in unique environments; one with shrimp as usual, one with crabs and a control with no prey items in the tank at all during the time of development within the egg.

The ones that were brought up in a tank with crabs would hunt and eat the crabs. The other 2 tanks would shun the introduced crabs in favor of the shrimp. To Jennifer Basil of Brooklyn City College, this may be evidence for behavior alteration in response to carefully observing their prey. Other scientist’s aren’t so sure and suggest that observational learning is restricted to the learning of a new behavior by watching a member of its own species; therefore behavior alteration due to being raised with crabs may be worth studying further, but not necessarily observational learning, at least in the eyes of some scientists.
An example in of this in humans – imagine you are looking for an Anatomy class, and you walk by two doors, one labeled ‘Anatomy’ the other ‘Chemistry’. If capable of reading the signs, the individual would obviously follow the direction of the signs to achieve their goal. While this is absolutely learning, its not an example of observational learning. An example of observational learning, which requires further cognitive processing than the former, would be a man at a foreign airport with unfamiliar language (where he couldn’t read M/F bathroom labels) figuring out which restroom he should use based on watching what other men were doing.

Octopods and other cephalopods are generally solitary animals, and an encounter between two octopods could be deadly for the smaller of the two. The use of changing color/texture is not only to hide; but also used for communication with other cephalopods. Displays called ‘passing cloud formations’, pictured above, are characterized by a series of dark lines running down the animals body to tips of its arms. It seems these displays are used to startle prey into moving while they remain still. The prey will be almost entranced by the ‘dancing’ patterns on the unique creatures skin, and as soon as it stops moving or comes closer, the octopus gains a meal.
The changing patterns aren’t always used to startle prey; If an octopus were placed in a tank with another octopus in a tank to the right, and a prey item in the tank on the left, it can divide its body into two halves; displaying a passing cloud formation towards the side of the prey, and a sort of tough-guy posturing towards the side with other octopus – as if to threaten not to take its food!

Examples of tool use in octopods are increasingly common, but still somewhat controversial. The evidence has been best summed up in two different examples. The first of which is of a tropical, pacific species of octopus, pictured above, which has been seen using coconut shells as a shield from predators; moving across the ocean floor covered by the coconut. Another example is a reef octopus species that has been observed building a rock wall up in front of its den to prevent the attack of a predator while the animal sleeps. This demonstrates at least a primitive ability to plan ahead; as the octopus builds up the wall in anticipation of predators when none are actually visible to the animal. While some scientists are reluctant to label this ‘tool use’ in the same way we refer to the ability of a chimp to modify a stick to ‘fish’ for termites; it is certain that these behaviors are indicative of an animal with a very complex and strategic cognitive process.
Why are Octopods interesting, Sexually?
Sex in octopods can be quite risky business, at least for the males. Female of several species of octopus, which are generally larger than the males, have been observed eating males who try to impregnate her. Males have adapted an absolutely amazing way of getting their genes in future generations, without causing their own death in doing so. Males have an arm (generally the third arm on the right side) called a hectocotylus. The hectocotylus is generally shorter and wider than the other arms, and the site of the male spermatophores. As is mentioned in the picture below, one means of sperm transmission occurs by the male octopus inserting his hectocotylus into the mantle of the female and releasing sperm; however, in species where female cannibalism is common, males have adapted a striking ability to ‘shoot’ the sperm from their hectocotylus at the female. Once launched, the sperm may travel several feet towards the female; ending near her arms where they burrow themselves in her skin. The male spermatophores may be stored for several months in the females skin and, when she is ready to reproduce, she can select the sperm (sometimes, that of multiple males) to father her many, many eggs. This allows males to transfer their sperm to the female without getting close enough to become prey themselves.

Octopods have notoriously short life spans, with dwarf species living an average of 6 months and the giant pacific octopus living maybe 5-7 years maximum. Females generally die after reproducing, and are actually quite exceptional mothers. After laying several hundred eggs in a carefully selected den, she will sit with them for several weeks while they develop. Octopods have a high metabolism, and need to eat frequently to survive under normal conditions. Hormonal influences occur following the females egg laying which decrease the animals metabolic rate; allowing the female to survive longer than normal without food. During the weeks spent in the den with her eggs, the mother starves herself to make sure predators dont make a snack of her babies while she goes hunting. The mother uses her arms to slowly caress the egg clusters. Causing this kind of disturbance in the water around her eggs helps to make sure her offspring have ample oxygen while developing. Generally, the hatching of the eggs is shortly followed by the death of the highly dedicated molluscan mother, who made the ultimate sacrifice for her offspring.

Why are Octopods interesting, Adaptively?
Octopod “Jelly-Bodies”- Octopods are notoriously difficult animals to keep in captivity, largely because of their ability to squeeze their entire bodies through the absolute smallest of openings. It is generally said that if the beak of the octopus could fit through a particular hole, so too could the rest of the animal. In marine aquariums, it is not uncommon for an octopod to escape the tank by climbing in the aquariums intake filtration nozzle, then following the series of tubes back to the main filtration system. Many octopods are capable of surviving for short periods of time on land as well. In fact, there have been several reports by aquarium hobbyists of their octopus climbing out of its tank (which was believed to be impenetrable) then crawling across the floor to climb in another one of their tanks to engorge itself on the fish and critters inside. Then, after eating its fill, the animal climbs out of the ravaged tank and slops along the floor back to its own tank; where it climbs back in through the external filter it exited through originally.
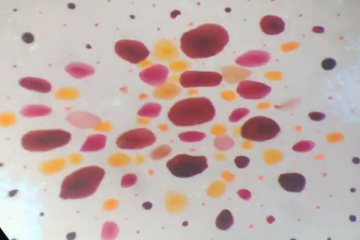
Octopod Chromatophores and Papillae: As was mentioned earlier, octopods don’t have a whole lot going on physically to deter predation by sharks, whales, seals and other marine predators. Instead, cephalopods rely on their ability to process large amounts of data about their environment very quickly and then immediately camouflaging themselves accordingly – in a manner that would surely make any chameleon jealous. They are able to match to both the color and the texture of their environment with near perfect accuracy using of sacs of pigment called ‘chromatophores’ (pictured above) and muscular regions called papillae (pictured below).

The best analogy I have heard of for a chromatophore is to think of one as a deflated balloon; the color of which alters as we tug at the ends of the balloon, thereby stretching and thinning the rubber material and allowing more light through. This analogy is limited, considering that if you used a blue balloon in this demonstration, you would only manage to get different shades of the original blue by stretching the material. However, I feel it still demonstrates a similar enough mechanism of color alteration as a result of the changing size of the pigment ‘sacs’. The expansion/contraction of octopod chromatophores dictates the color of that particular region of the animal.
The color of human skin is dictated by multiple genetic and developmental determinants that influence the activity of melanocytes, which create the pigment melanin. The more active ones melanocytes are, the more melanin they will produce; the more melanin they produce, the darker their skin will be. Humans and other vertebrates are largely unable to change the color of their skin. Certainly not as quickly and drastically as octopods, whose distributed neural ganglia collaborate with the animals chromatophores to alter the color patterns as needed.
Papillae are protrusions off the octopods skin that are formed by underlying muscle forcing up folds of skin. These folds are extremely similar in appearance to the algal growth common on rocks in their marine scape. The octopus is able to look at its environment and very quickly assess the best way to camouflage its body from marine predators; if it is in a region where algae is common, it would almost certainly utilize its papillae to mimic the texture of the surrounding algae and its chromatophores to mimic the color of its surrounding. This, in effect, renders the animal invisible to its pursuant, and allows the cephalopod to see another day.
Just as any proper Darwinian would expect, the amount of chromatophores present in the skin of the cephalopod depend largely on the region from which the animal is taken. For example, octopods living in coral reefs where there is an abundance of light and predators that use their eyesight to locate their prey; chromatophores are abundant in the animals skin. By contrast, Cephalopods living deep in the ocean where light does not penetrate have very few, if any, chromatophores – as they would not have survival benefit where predators do not rely on eyesight and the presence of light to hunt.

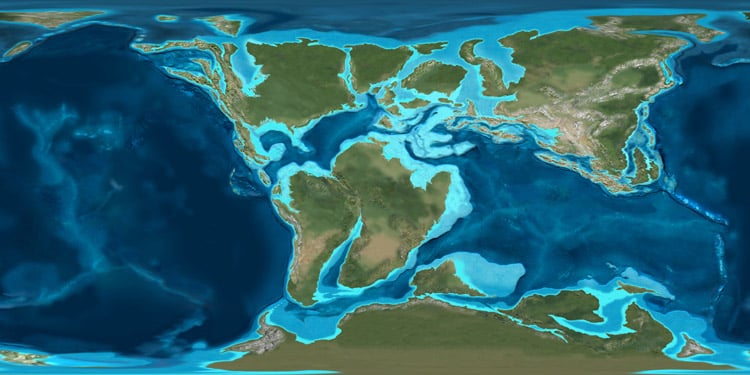
Octopod Distribution: Octopods have a marine distribution that any other sea creature would be jealous of. Found in waters both frigid and tropical, from the shallow coasts to the deepest depths science has been able to explore. Chances are, if you are near an ocean, a cephalopod of one kind or another is not far. The real question is whether or not you can see them. Their fascinating adaptations have allowed them to be successful predators of the shallows and the extreme depths. I suspect that as science advances, allowing us to explore even deeper in our oceans, we will continue to find cephalopods living much as they have been for hundreds of millions of years. Basically, in the above picture, cephalopod territory is marked in blue. 🙂
*More information on my (soon to begin) cephalopod research and, of course, the work I have been doing with Bubba will be up ASAP.
References:
Tamar Gutnick, Ruth A. Byrne, Binyamin Hochner, Michael Kuba. Octopus vulgaris Uses Visual Information to Determine the Location of Its Arm. Current Biology, 2011; 21 (6):
Hochner, Binyamin. “Neurobiology Talk tomorrow – Binyamin Hochner on Octopus Neurobiology.”Neuron Club. (2012): n. page. Web. 12 Apr. 2013. <http://neuronclub.org/2012/10/17/neurobiology-talk-tomorrow-binyamin-hochner-on-octopus-neurobiology/>.
Hochner, B., Shomrat, T. and Fiorito, G. (2006). The octopus: a model for a
comparative analysis of the evolution of learning and memory mechanisms. Biol.
Bull. 210, 308-317
Mather, J. A. (1995). Cognition in cephalopods. Adv. Study Behav. 24, 317-353.
Hanlon, R. T. and Messenger, J. B. (1996). Cephalopod Behavior. Cambridge:
Cambridge University Press.
Boing Boing Science – (https://www.youtube.com/watch?v=KyGazPZmmM0)
Photo References:
Cephalopods pic (at top) – http://tolweb.org/Cephalopods
Micro-Mollusks pic- http://upload.wikimedia.org/wikipedia/commons/thumb/f/ff/Mudsnail2.jpg/220px-Mudsnail2.jpg
Phylum Mollusca Pic- http://www.britannica.com.ph/article_images/72197.jpg
Giant Squid Dissection: http://museumvictoria.com.au/pages/6981/gallery/08-0281-188.jpg
Octopus Hectocotylus-http://www.asnailsodyssey.com/IMAGES/OCTOPUS/octopusMate.gifReef Camouflage Pic- http://www.mentalfloss.com/sites/default/legacy/wp-content/uploads/2008/09/434camouflage.jpg
Chromatophores-http://www.technewsdaily.com/images/i/000/007/714/iFF/squid-chromatophores.jpg?1348673396
blue ring/papillae- http://24.media.tumblr.com/tumblr_lfw6xq55qw1qessr0o1_400.jpg
human v octo eye- http://thegenomestale.files.wordpress.com/2012/05/f1-large.jpg
octopus suckers- http://tvblogs.nationalgeographic.com/files/2011/12/3309a2dc-8f99-49bf-87f9-6b7ed70fa6f5.Full_.jpg
Octopus beak- http://www.nwboat.com/pictures/beak.jpg
Octopus in coconut- http://images.mnn.com/sites/default/files/octopus-builds-shell.jpg
Radula Drawing- http://www.asnailsodyssey.com/IMAGES/CHITON/radulaDrawing.gi
Real Radula- http://depts.washington.edu/fhl/zoo432/floats/flfeeding/Katharina_radula.jpg
Mom and Eggs- http://cdn1.arkive.org/media/50/50D69813-79E7-4A60-B4EC-85E3671677C0/Presentation.Large/Female-North-Pacific-giant-octopus-guarding-eggs.jpg
Snail anatomy- http://images-mediawiki-sites.thefullwiki.org/03/1/6/7/8798767243165146.png
Octopus anatomy/buccal- http://jeb.biologists.org/content/210/1/118/F1.large.jp
Octopus papillae-http://www.divephotoguide.com/images/imguploader/948520026.jpg
Octopus anatomy/hearts: http://www.canaryzoo.com/Around%20World%20QA/Octopus%20Hearts.jpg
Aristotle: http://4.bp.blogspot.com/_wuSqJG5bIKE/TB_Y_Og2QpI/AAAAAAAAAhk/I75qMP2HbxU/s1600/Aristotle.jpg
Invert vs Vertebrate Neurons- http://vanat.cvm.umn.edu/neurLab1/images/Lab1Fg01Opt.gif
Cuttlefish- http://www.globefish.org/upl/Cephalopods/cuttlefish.jpg












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}